Relationships have been established between dermatoglyph profiles and temperament (Grandin et al, 1996; Randle 1998; Nash, 2006; Gorecka et al, 2006; Olmos et al, 2008), laterality (Murphy and Arkins, 2008; Randle et al, 2010) and abnormal behaviour (Swinker et al, 1994; Luescher et al, 1991; van Oel et al, 2001; Gourion et al, 2004; Hale et al, 2005) in multiple species. Within the equine industry, the performance of stereotypic behaviours (SBs) is considered to impair performance and reduce value in affected individuals (Winskill et al, 1995; Mills et al, 2002), potentially reducing career longevity and compromising welfare through the deployment of strategies to prevent their expression when they are established (McGee, 2006).
The ethological and physiological arguments made within previous research regarding the aetiology of SBs differ. SBs have been defined as functionless repetitive behaviours which incorporate flexible motor patterns (behaviour is goal directed with each repetition) (Mills and Luescher, 2006). In contrast, Mason et al (2007) proposed that stereotypies are coping mechanisms which may relate to the ontogeny of repetitive behaviour. Garner (2005) broadened the definition identifying ‘abnormal repetitive behaviour’ (ARB) as behaviours where it was impossible to ascertain the cause, and/or where the negative impact on welfare or natural frequency was unknown (the definition utilized for the current study). In the horse, locomotor (for example weaving), comfort/oral (for example crib biting) and social (for example head circling) stereotypies have been associated with social isolation in horses (Mills et al, 2002) although Dodman et al (2005) have suggested that not every horse exposed to stressful environments develops SBs or indeed ARBs.
In both humans (Tanimura et al, 2008) and equidae (McBride and Hemmings, 2005) SBs are associated with alterations to the cortico-basal ganglia (CBG) circuitry. Dopamine is thought to be the primary neurotransmitter involved in the expression of SB with the ventral striatum recognized as the primary neural centre for goal-directed behaviour (McBride and Hemmings, 2009) and to produce enhanced motivational states in stimulus-response learning in affected individuals. Thoroughbreds exhibit a higher percentage of SBs compared with other breeds (Waters et al, 2002; Bachmann et al, 2003; Albright et al, 2009); with both Lloyd et al (2008) and McBride and Hemmings (2005) demonstrating that thoroughbreds are more reactive to stimuli than other breeds. This may not be surprising as modern racehorses represent a limited population with selection based on performance rather than temperament or capacity for environmental adaptation (McGee, 2006; Thiruvenkadan et al, 2009).
Dermatoglyphics is the study of dermal directional patterning and considers location and morphological features of trichoglyphs (whorls) (Smith and Gong, 1973). The influence of intra-uterine epigenetic factors during early gestation has been proposed as a mechanism that drives dermatoglyph profiling in the human foetus (van Oel et al, 2001; Youseti-Nooraie and Martaz-Hedjri, 2008) and bovine foetus (Olmos and Turner, 2008). Embryogenesis of cerebral, neural and dermal tissue all originate from the ectodermal layer of the foetus (Paine et al, 2003) and dermal patterning is considered to be an external indicator of cerebral architecture of the individual (Smith and Gong 1973, 1974; Swinker et al, 1994; van Oel et al, 2001).
Relationships between dermatoglyphic profiles (scalp whorls) and cerebral abnormalities have been established in humans, for example schizophrenia (Alexandra et al, 1992; van Oel et al, 2001; Gourion et al, 2004;) and obsessive compulsive disorder (Luescher et al, 1991). Scalp whorls are considered to provide an external indicator of phenotypic expression of genes linked to neuro-cognitive function. To date research in the horse has linked trichoglyph position to temperament (Gorecka et al, 2006) and laterality (Elworthy, 2004; Murphy and Arkins, 2008; Randle et al, 2010).
No previous research has examined the links between equine dermatoglyphic profiles and expression of equine ARBs. A mechanism that could identify individuals susceptible to SB or ARB development from birth could be utilized to reduce environmental stress via deployment of prophylactic management practices to improve welfare or potentially performance.
Materials and methods
The current study investigated the relationship between dermatoglyphs and behaviours described as ARB, including those behaviours traditionally referred to as SBs. Behaviours were considered abnormal if the frequency of expression exceeded three repetitions within a 30 minute time frame with no apparent motivation (McDonnell, 2008), as this could potentially impair physical or psychological health. ARBs included in analysis demonstrate the potential for reduced performance, for example door kicking (front feet) may induce repetitive strain injury which could be influential in the high level of superficial digital tendonitis exhibited within racehorses (Williams et al, 2001). Behaviours were categorized into locomotor (Lo), social (S) or oral (O) origin (Table 1).
| Locomotor | Oral/comfort/eating | Social |
|---|---|---|
|
|
|
|
The sample comprised thoroughbred horses (Equus caballus) (n=41: 68% geldings and 32% mares) actively engaged in training for racing (Yard 1 jump racing, n=24; and Yard 2 fat racing, n=17) of variable age (> 1 year; mean age of 6.39±2.47 years, range 3–13 years), and height and colour. Expression of ARBs were ascertained via trainer interviews. At the commencement of each interview trainers were presented with a sheet defining ARBs to be assessed; trainers then identified actual ARBs that were performed, and at what frequency, by individual horses. Observations of selected subgroups within each yard were employed to test the validity of the data provided by the trainer. Individual horses were observed unobtrusively for 30 minute periods over three occasions on the same day by a constant observer: prior to feeding; prior to exercise; and when stabled post exercise (an hour after food with no forage present). Husbandry practices (diet composition and feeding frequency, turnout, exercise, social contact) were also recorded to enable consideration of environmental stimuli on behaviour. Bloodlines were not considered as the gene pool of the modern thoroughbred is known to be limited, suggesting heritability of phenotypic traits amongst all individuals is high (Thiruvenkadan et al, 2009).
A dermatoglyph profile was produced for each horse using previously validated methods (Nash, 2006; Murphy and Arkins, 2008) but expanded to include a full body analysis (i.e. not just facial pro-file) and recorded on a standard Royal College of Veterinary Surgeons' equine identification form. Individual trichoglyphs were photographed, by the same experimenter, using a 10.2 mega-pixel Canon 400D digital camera. A tape rule was placed parallel to the dorso-ventral orientation of the whorl to avoid variance between individual horses/whorls during photography and enable accurate measurement during subsequent remote computer analysis (using Adobe Photoshop: version 9.2).
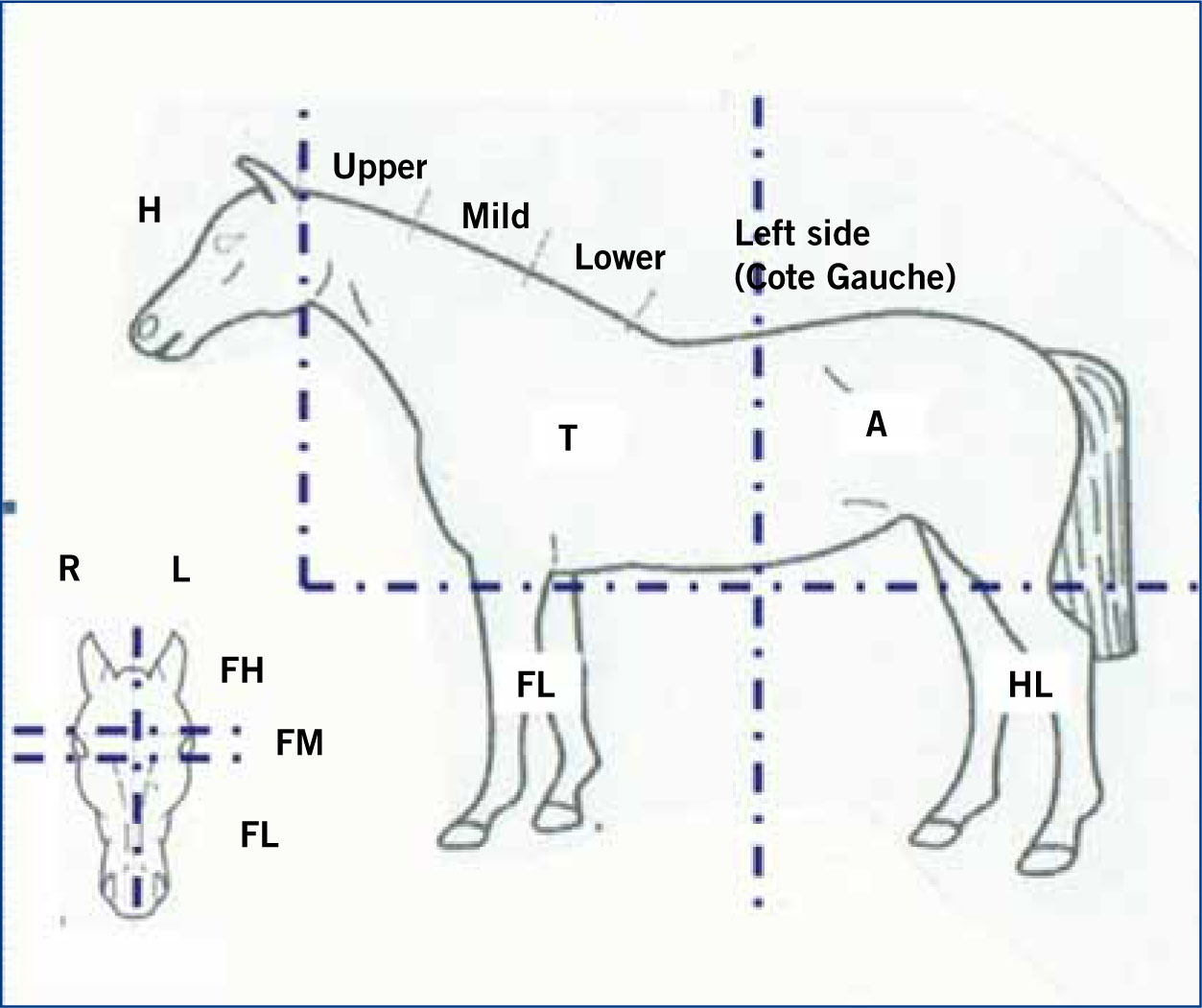
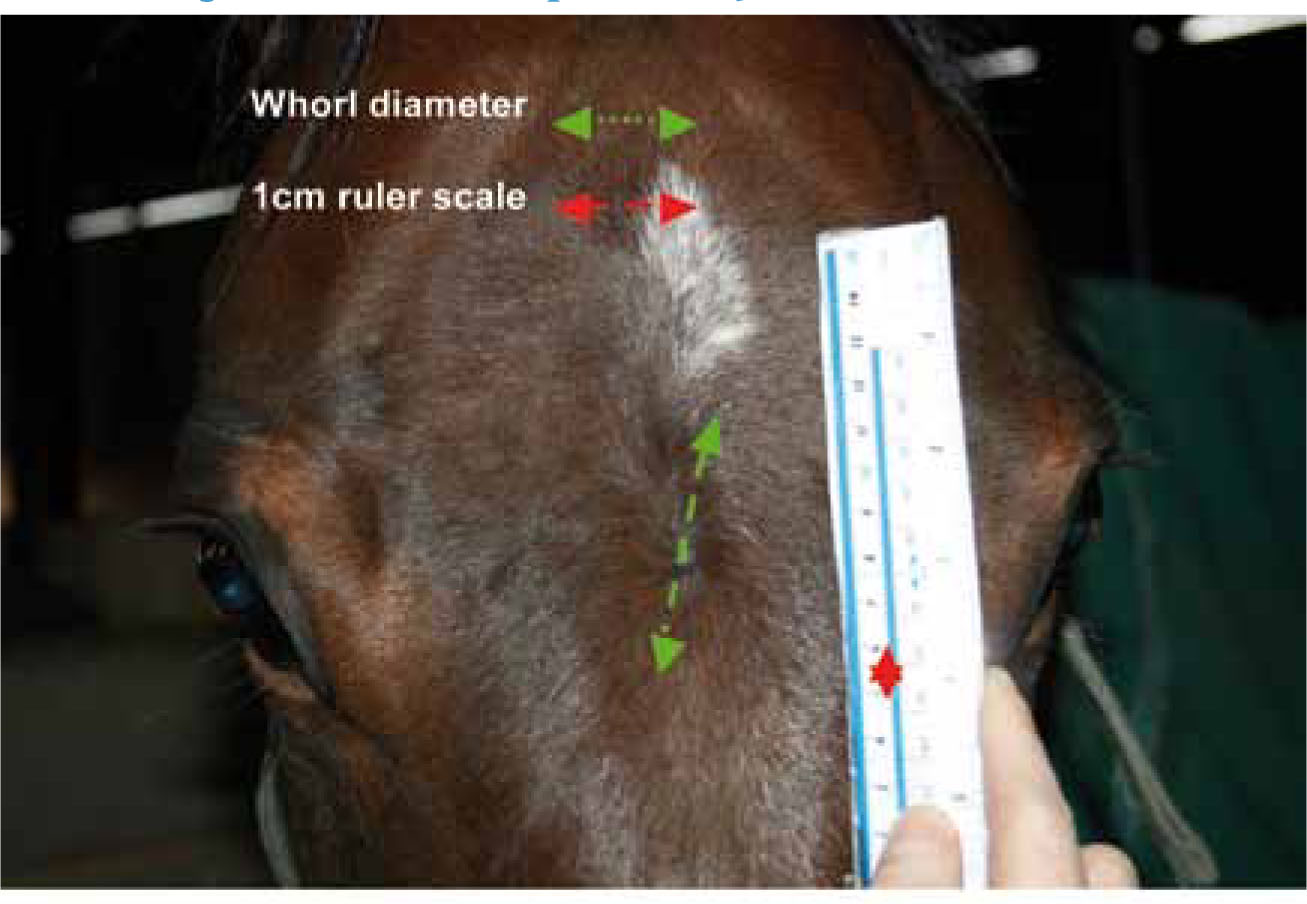
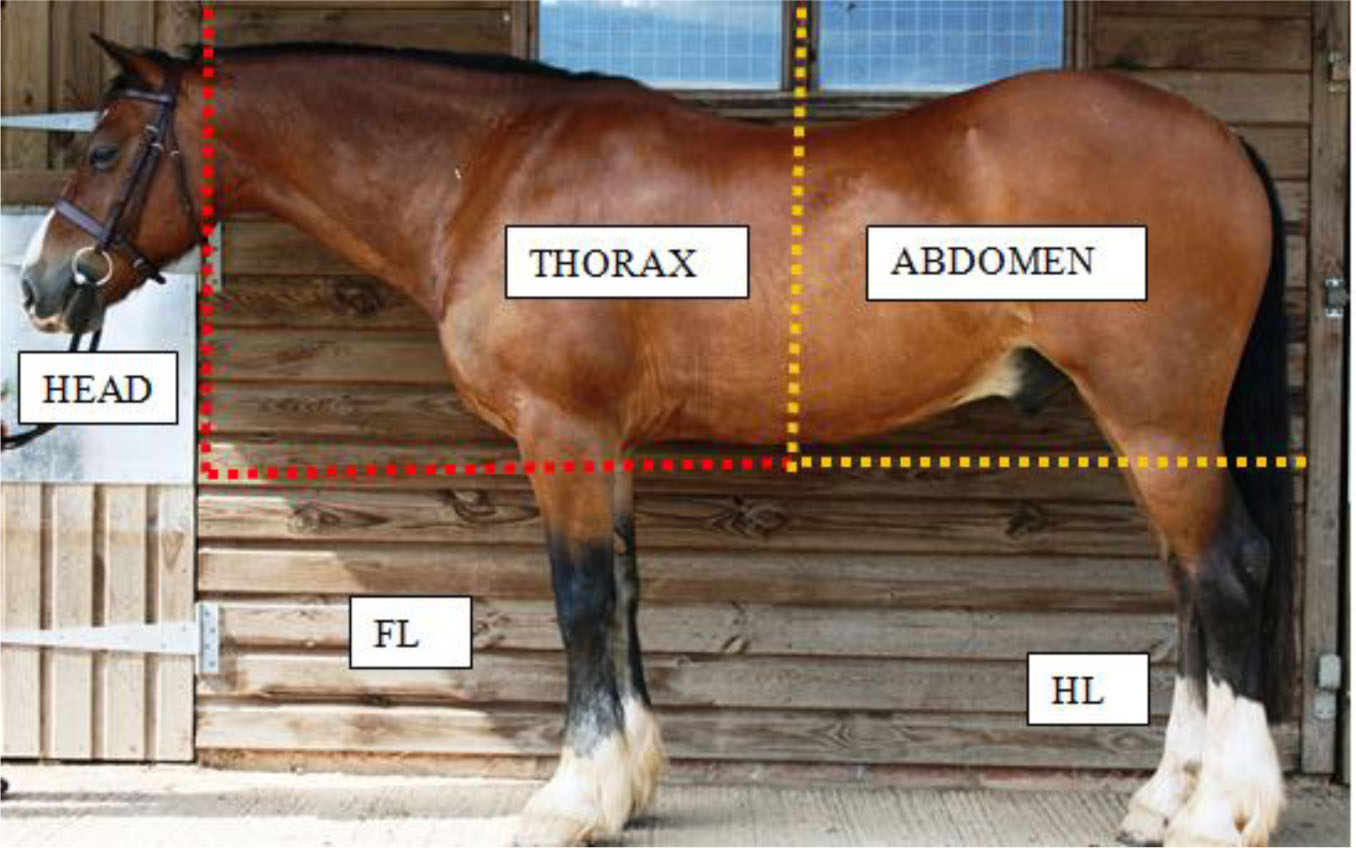
For the purpose of data collection and analysis the horse was divided into segments (Figure 1). The total number of whorls for each segment and the entire horse was noted with any double (side by side) or paired whorls (per body segment) identified. Trichoglyph orientation was identified as clockwise (C), counter-clockwise (CC) or radial (R) as used by Grandin et al, (1996), Randle (1998) and Murphy and Arkins (2008). Whorl morphology was recorded with reference to whorl shape: round (WR) or elliptical (WE), maximum diameter of trichoglyph (MDT) and epicentre shape: round (ER) or not round (ENR). MDT, a novel parameter, was defined as the distance between the borders of directional hair patterning incurred as a specific outcome of whorl hair follicular growth (Figure 2). A comparison of trichoglyph positional (2D/segment) and orientation symmetry/asymmetry (3D/lateral placement) was conducted in reference to the right and left side of the horse for all segments under investigation (Figure 3a and b).

Data were analyzed using non-parametric statistical tests. Associations between SB and ARB expression and gender, training establishment, trichoglyph morphology and location were tested using Mann-Whitney U-tests. A series of Kruskal-Wallis Analysis of Variance were conducted to compare ARB expression and colour and trichoglyph morphology. Spearman Rank Correlation Co-efficient tests were used to test for correlations between location, orientation and number of trichoglyphs and expression of ARBs. Environmental impact on expression of stereotypies was evaluated by comparison of expression of the total number of ARBs between Yard 1 and Yard 2. Additionally a series of Mann Whitney U analyses for individual, categories and total number of stereotypical behaviours observed within the two sample populations were conducted.
Ethical approval for the study was granted by the University of the West of England Ethics Committee.
Results
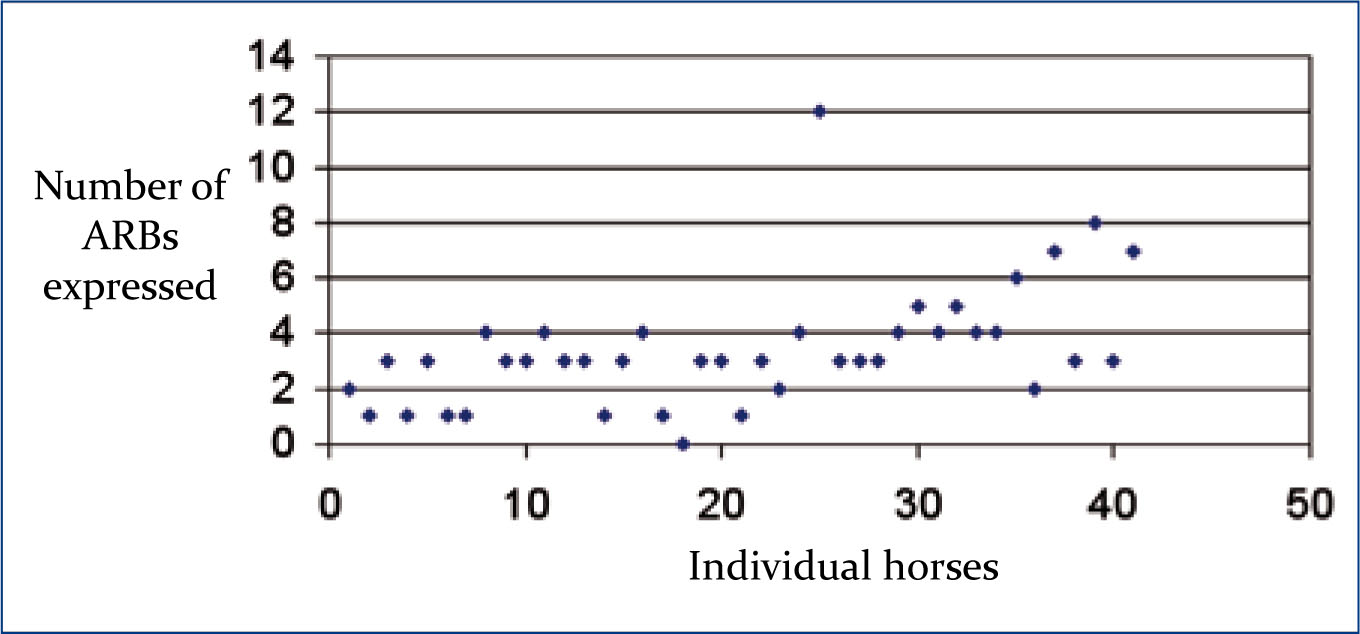
The majority of the sample population (97.54%) exhibited ARBs, with a mean of 3.41±2.21 sum ARBs expressed per individual horse. Figure 4 illustrates the total number of ARBs exhibited per horse. In the current study 28% of horses performed social ARBs, 15% exhibited locomotor ARBs, and 97% performed oral ARBs. Of the whorls examined, 305 (92%) were elliptical and 27 (8%) were round in shape, whilsethe epicentre shape was recorded as 314 (94.5%) not round and 18 (5.5%) round. Each individual in the study presented with whorls with a mean orientation of CC = 3.54±1.73; C=3.00±1.94; R=1.83±1.67 for the whole body and CC = 0.57±0.84; C = 0.44±0.63; R = 1.15±0.79 for the face. Within the sample 61% (n=54) of facial trichoglyphs were located above the eyes, 30% (n=26), were located between the eye lines and 9% (n=7) were below the eyes. Significant relationships exist between whorl morphology and expression of ARBs (Table 2).
| Parameters | Result |
|---|---|
| Box walking: number of abdominal whorls | Spearman's rho = 0.34; p<0.05* |
| Head nodding: number of abdominal whorls | Spearman's rho = 0.35, p<0.05* |
| Expression social ARBs: number of abdominal whorls | Spearman's rho = 0.40, p<0.01** |
| ER: expression of O ARBs | Mann Whitney U = 53024, p=0.052* |
| ER: sum expression of ARBs | Mann Whitney U = 53299, p<0.05* |
| WR: head nodding | Mann Whitney U = 51761.5, p<0.05* |
| WR: expression of Lo ARBs | Mann Whitney U = 51364.5, p<0.05* |
| WR: expression of S ARBs | Mann Whitney U = 52224, p<0.001† |
| WR: sum expression of ARBs | Mann Whitney U = 52614, p<0.0001†† |
| Thorax CC: head nodding | Spearman's rho = 0.35, p<0.05* |
| Abdominal CC: lip licking | Spearman's rho = − 0.36, p<0.05* |
| Abdominal C: head circling | Spearman's rho = − 0.34, p<0.05* |
| Abdominal C: expression of S ARBs | Spearman's rho = 0.32, p<0.05* |
| Abdominal C: sum expression of ARBs | Spearman's rho = −0.39, p<0.05* |
| Single facial R: expression of O ARBs | Kruskal-Wallis H4=8.89, DF = 2, p<0.05* |
ARB, abnormal repetitive behaviours; O, oral; L, locomotory; S, social; CC, counter-clockwise; C, clockwise; R, radial;
Comparison of exhibition of ARBs between the two training establishments found that horses in Yard 2 demonstrated significantly more individual ARBs than Yard 1 than expected by chance for all variables that could be analyzed (chewing: W=428, p<0.05; wood biting: W= 569, p<0.05; crib biting: W=449, p<0.05; wind sucking: W=449, p<0.05; weave side-to-side: W=452, p<0.05; weave head swing: W=452, p<0.05; head nod: W=418, p<0.01). The incidence of oral ARBs performed at Yard 1 was 17% lower than that of Yard 2. Similarly the percentage of locomotor ARBs and social ARBs were lower in Yard 1 than Yard 2 (79% lower Lo and 53% lower S).
Discussion
The results of this study suggest dermatoglyphic profiles may be an external representation of phenotypic predisposition to perform ARBs in the horse. Differences appear to exist between the ontogeny of oral ARBs and locomotor and social ARBs, implied by the lack of strong trends in the latter category. The majority of the sample presented with whorls with elliptical gross and epicentre morphology, while analysis suggests ‘abnormal’ trichoglyph morphology (round gross and round epicentre morphology) increases the likelihood of ARB expression in Lo and S categories. Meola et al (2004) previously correlated round facial whorl morphology to abnormal semen motility and morphology in Angus bulls. Interestingly testicular development occurs at the same time as notochord and epidermal tissue during embryogenesis; these results suggest that whorls may provide a physical clue to physiological or biochemical stressors that have occurred during embryogenesis.
Few correlations were found between whorl location and morphology with the performance of ARBs, however abdominal whorl orientation asymmetry corresponded to increased performance of ARBs. Bilateral symmetry of physical attributes is considered to externalize an individual's phenotypic quality to respond to pre-natal environmental insults during foetal development (McManus, 2002; Fink et al, 2006). Environmental stressors that occur within the uterine environment during differentiation of the ectoderm may subsequently be recorded via dermal patterning (whorl) asymmetry and provide external clues to underlying neurological dysfunction.
It appears that MDT is inversely proportional to an individual's predisposition to express abnormal behaviour; with horses with small, tight whorls being more likely to performs ARBs. It could be suggested that thoroughbreds with large MDTs would be less likely to exhibit ARBs, and could therefore react inherently less to environmental challenge. The selection of these individuals to engage with racing could reduce perceived negative welfare aspects of management regimens; it would also be interesting to evaluate the relationship of this parameter to performance ability.
Interestingly, horses within Yard 2, fat racing, exhibited a higher percentage of all categories of stereotypy than horses in Yard 1, jump racing. Thoroughbreds engaged in jump racing are often products of an unproductive fat career (25% of jumpers in this study had recorded performance ratings on the fat) therefore breeding lines do not necessarily differ (McGee, 2006). Exercise, turnout and feeding regimens were comparable, however Yard 1 did provide environmental enrichment (gorse bush branches) but the main differential observed was housing. Yard 1 was of American Barn style construction with half-height partitions between stalls to purposefully facilitate social contact, in contrast to Yard 2 which was of traditional brick built stables with frontal door and closed window facing onto a central courtyard. Previous work has demonstrated that restricting social behaviour increases development of stereotypies (McGreevy et al, 1995a; Waters et al, 2002; Nagy et al, 2008). The reduced opportunity for social interaction at Yard 2 could expose horses to increased environmental stressors and correspond to the enhanced demonstration of stereotypies observed.
A number of individual ARBs were not performed by enough individuals in this study and therefore statistical analyses were not possible (due to very small and/or equality of dependent variables or zero occurrences of individual ARBs). Further research incorporating increased numbers of horses and training establishments are required to substantiate these findings.
Horses in the current study demonstrate increased performance of ARBs compared with research previously conducted (Table 3). However, it should be noted that previous work has concentrated solely on SBs and does not include the potential expression or impact of ARBs. Thoroughbreds represent a closed population, arising from four foundation sires and seven foundation mares (Thiruvenkadan et al, 2009). This combined with the deployment of similar management practices (typically abrupt weaning, limited turnout, low forage: high concentrate diets, limited social contact) appears to provide unique opportunities for the enhanced expression of ARBs in comparison to other breeds.
| Stereotypy | Reported incidence (%) | Reference |
|---|---|---|
| Total abnormal behaviour |
|
|
| Weaving | ||
| Crib biting | ||
| Box walking |
|
|
| Wood chewing |
|
Specifc to thoroughbred bloodlines;
within thoroughbred populations exclusively;
within 2 year olds stabled in individual stalls for the first time.
Sixty one percent of thoroughbreds in this study had high facial whorls. Research in both cattle (Tanner et al, 1994; Grandin et al, 1996; Randle, 1998; Olmos and Turner, 2008) and horses (Gorecka et al, 2006; Nash, 2006) has found that individuals with high facial whorls are more behaviourally reactive than their counterparts. The results support Lloyd et al's (2008) finding that proposed thoroughbreds are highly reactive in comparison with other breeds. It could be suggested that an inheritance factor (increased CBG sensitivity) produces these high reactors which are greater in number in the thoroughbred and predisposes affected individuals to over react to reduced environmental cues (McBride and Hemmings, 2009).
The relationship between enhanced learning capacity and highly emotive environments has been established in numerous species (Dodman et al, 2005). For individuals who exhibit phenotypic predisposition to perform ARBs (due to increased CBG sensitivity) stronger reinforcement and enhanced neural plasticity could increase the likelihood of them developing and exhibiting increased ARBs as they age (Luescher et al, 1998; Dodman et al, 2005; Nicol, 2005). Further research is required to identify whether inheritance of dermatoglyph profiles (whorls) influences performance of ARBs in of-spring with consideration of the short and long-term impact of environmental variables in the study design.
Conclusion
The thoroughbred population examined exhibited higher than expected frequencies of ARBs. The evidence presented in this study indicates that dermatoglyphic profiles are a viable external representation of phenotypic predisposition to perform ARBs in the thoroughbred. The racing industry could utilize this information to identify individuals predisposed to ARBs and adapt their management regimens to prevent their development thus retaining their economic value and preventing related injury. Dermatoglyphic profiles (whorls) exhibit potential for the improvement of horse welfare, via informed application of management practices, if the dermatoglyphic patterns found in the thoroughbred can reliably predict highly reactive individuals within other breeds. Identification of susceptible individuals would allow management of environmental cues and stressors hopefully to reduce the negative impact of ARB expression on performance (e.g. development of laterality bias in racehorses) or to facilitate their expression as a component of a coping mechanism.